在动物和植物中,H3K9me3和H3K27me3染色质沉默标记通过不同的蛋白质机制被留存下来 。 H3K9me3是由SET-domain SU(VAR) 3-9酶催化的,而H3K27me3是由集域增强zeste酶催化的, 它们都是Polycomb Complex 2 (PRC2)的催化亚基。此处研究者发现来自单细胞真核草履虫的zesteer -like蛋白Ezl1,与人类EZH2基因序列和结构相似,在体内外催化组蛋白H3的甲基化,具有明显的K9和K27位点特异性。同时发现H3K9me3和H3K27me3同时出现在多个家族中,证明了这些损失组蛋白标记导致转座因子的整体转录超活化对蛋白质编码基因表达的适度影响。
我们的研究表明,尽管经常如此H3K9me3和H3K27me3在功能上是不同的,它们可能有一个共同的进化过程历史以及在沉默转座因子方面的共同祖先作用。
组蛋白的翻译后修饰在染色质水平的DNA事件中起着重要作用。在这些修饰中,组蛋白H3在赖氨酸9 (H3K9me3)和赖氨酸27 (H3K27me3)上的三甲基化通常与转录沉默的染色质有关,这两个表观遗传特征由组蛋白甲基转移酶的不同家族控制。人们普遍认为H3K9me3和H3K27me3并不在同一个基因组位点上协同工作,这一假设得到了一些生物体证据的支持。在人和果蝇中,H3K9me3富集域主要集中在 重复的DNA序列,如转座因子和卫星,抑制潜在重复的转录。相比之下,H3K27me3参与了细胞型特异性基因抑制的稳定和遗传维持。
然而,H3K9me3和H3K27me3同时出现的例子对这一观点提出了挑战。事实上,ChIP-seq数据表明,这两个标记可以同时在小鼠胚胎干细胞、胚胎外谱系和人类分化的发育调控基因子集中发现 细胞。此外,蛋白鉴定数据表明,H3K27me3可以与H3K9me3在胚胎干细胞的同一组蛋白八聚体上共存。
Ezl1 targets methylation of both K9 and K27 of histone H3.
Ezl1靶向组蛋白H3的K9和K27的甲基化 通过与野生型H3或单突变体(K9R和K27R)相比,在K9R/K27R双突变体H3上检测到甲基转移酶活性降低(图2d),这表明Ezl1也针对其同源H3底物中的赖氨酸9和27,H526A突变也严重影响了Ezl1的甲基转移酶活性。
通过使用Masquant软件对数据进行分析比对,有135个磷酸化位点是较为可信的。 一般来说在质谱峰图中可以在低质量区域看到亚胺离子的很强的信号,会看到m/z为190.037的离子峰的磷酸化组蛋白肽段,如图2b.
为了进一步研究草履虫H3中哪些赖氨酸残基以ezl1依赖的方式甲基化,我们进行了定量无标记质谱分析。 体外甲基化反应用含有草履虫组蛋白H3的四聚体和Flag-FA双亲和纯化的Ezl1(wt)和Ezl1(H526A),或仅用四聚体作为阴性对照。在此条件下,Ezl1催化K9和K27的三甲基化(图2e,表1,补充数据1)。 我们可以蛋白质谱定性到组蛋白H3 的N端存在其他甲基化位点,尤其是K23,但在催化Ezl1H526A突变体中,只有K9和K27上的甲基化显著降低(表1,图2e)。

表1
检测其他赖氨酸甲基化解释了剩余赖氨酸9和27突变时, 发现在非洲爪蟾蜍或草履虫H3(图2 c, d)。使用蛋白定性质谱分析和K9R / K27R 双突变草履虫组蛋白H3,研究者的确发现K23甲基化位点当赖氨酸9和27不再是酶的底物时候通过依赖蛋白Ezl1发生修饰。总之,所有试验数据支持了赖氨酸9和27都是Ezl1的主要底物的假设,而K23是Ezl1的次要底物,甲基化在体内的相关性需要进一步研究。
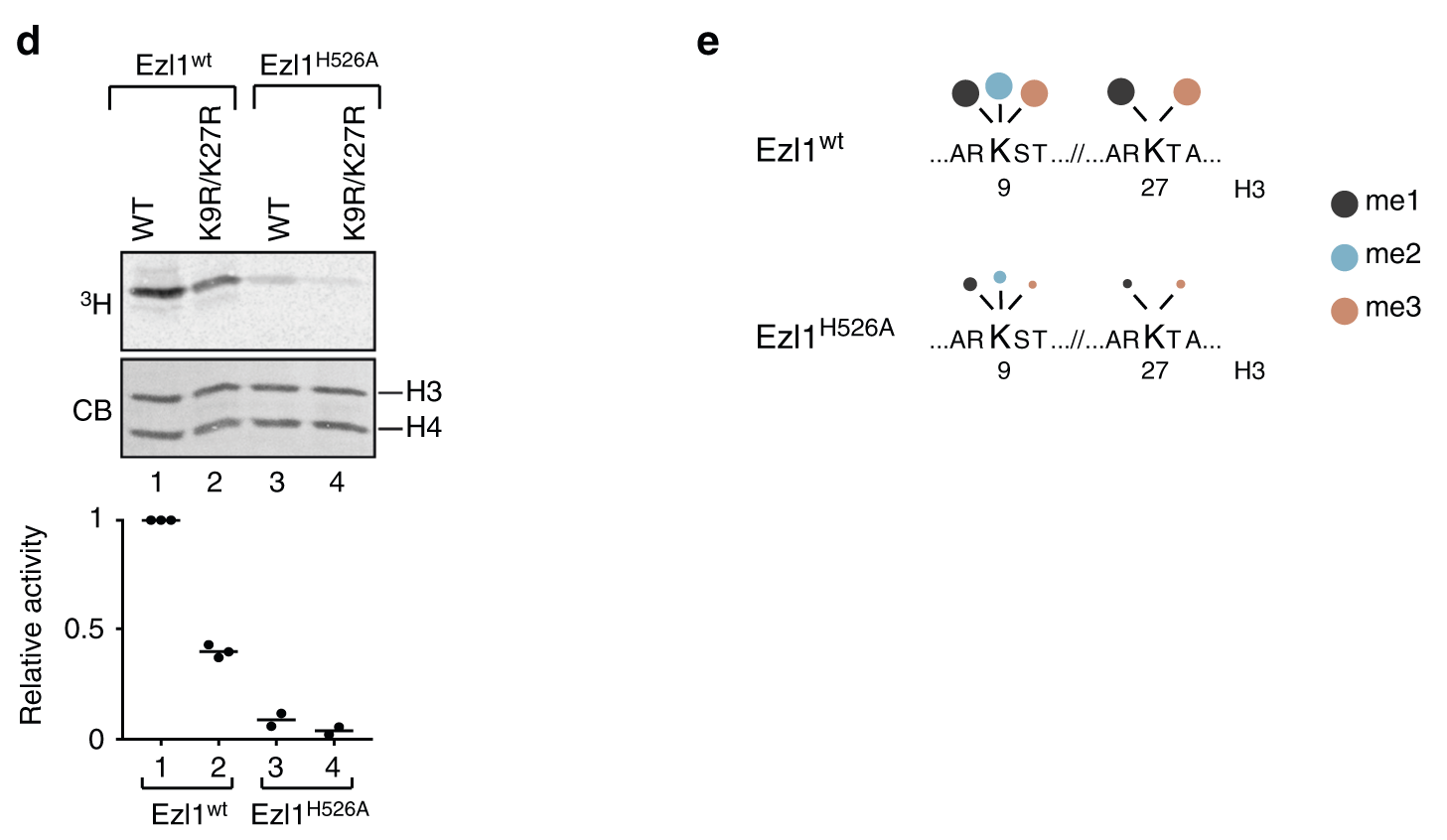
图2
为了研究Ezl1在体内的催化活性,作者又进行了基因互补在内源性EZL1基因被敲除的细胞中,表达野生型或突变型的GFP-EZL1转基因,进行检测。在对照条件和EZL1 RNAi条件下,GFP融合蛋白均定位于转化细胞的受精卵MAC中,表明每个RNAi耐药转基因均产生具有正确核定位的融合蛋白。 此研究过程中通过蛋白质谱手段确定了组蛋白的甲基化位点,又通过其他分子生物学手段找到相关试验证据,可见质谱对特定蛋白修饰位点鉴定已成为各研究者的必备手段,AIMS可以提供组蛋白的甲基化位点鉴定服务。
参考文献:SARS coronavirus protein nsp1 disrupts localization of Nup93 from the nuclear pore complex
Aimsmass---蛋白质谱服务商,提供快速便捷的组蛋白的甲基化位点鉴定服务


