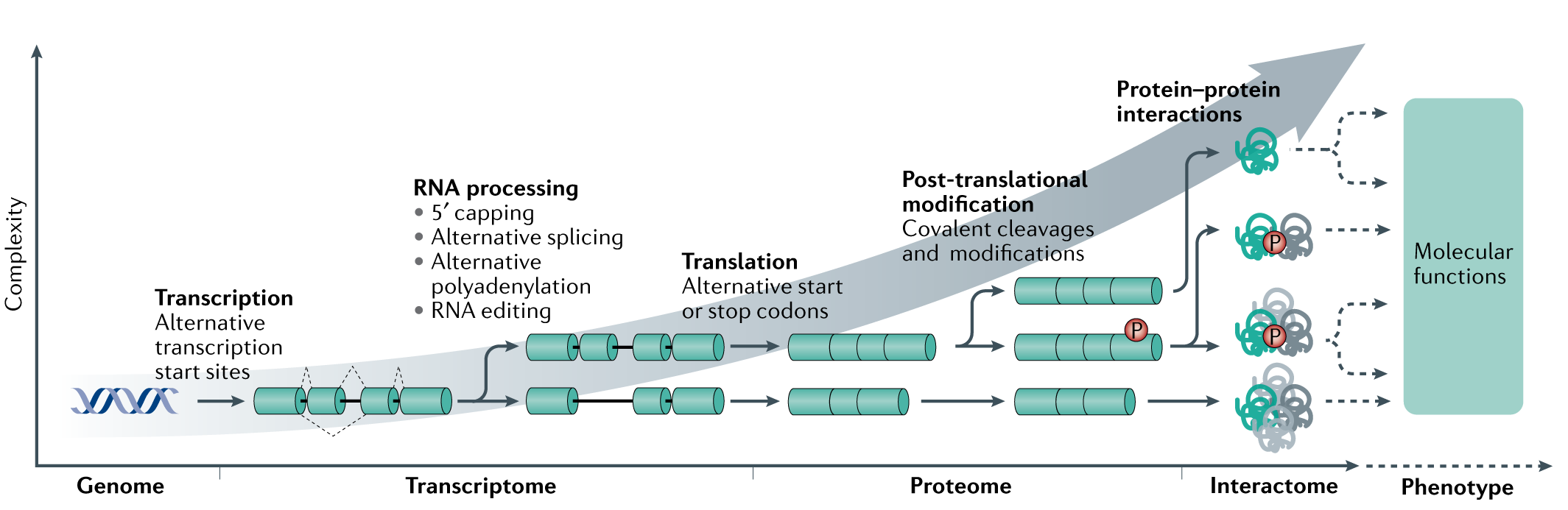
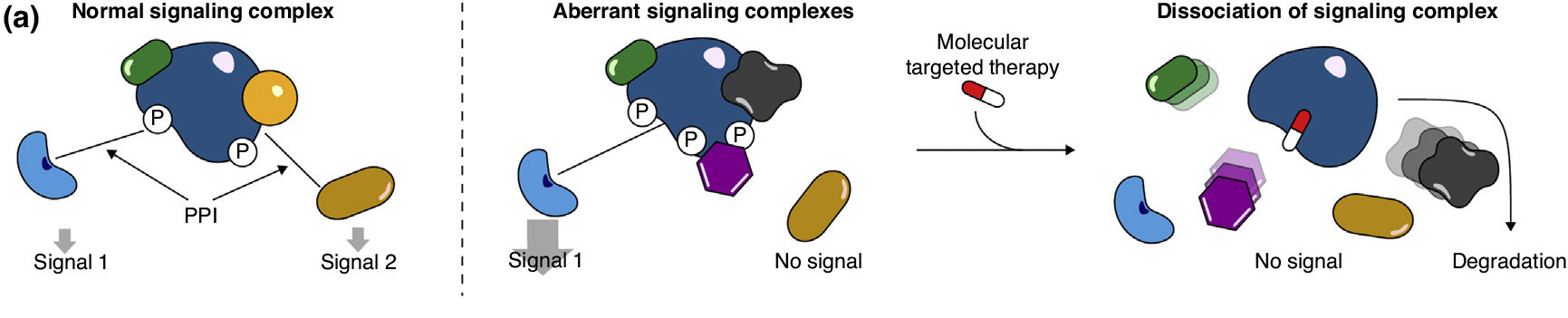
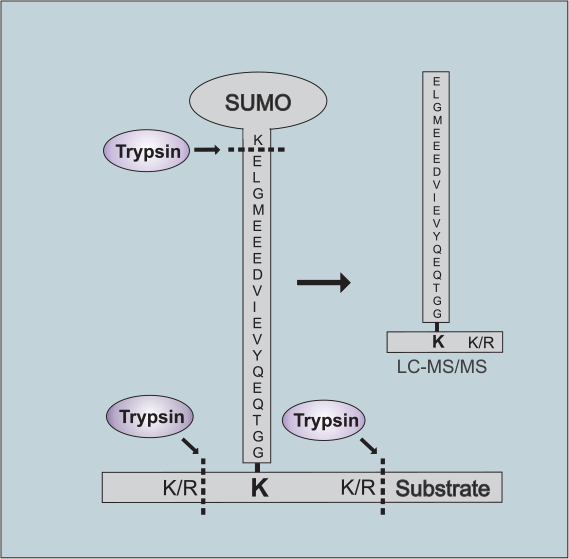
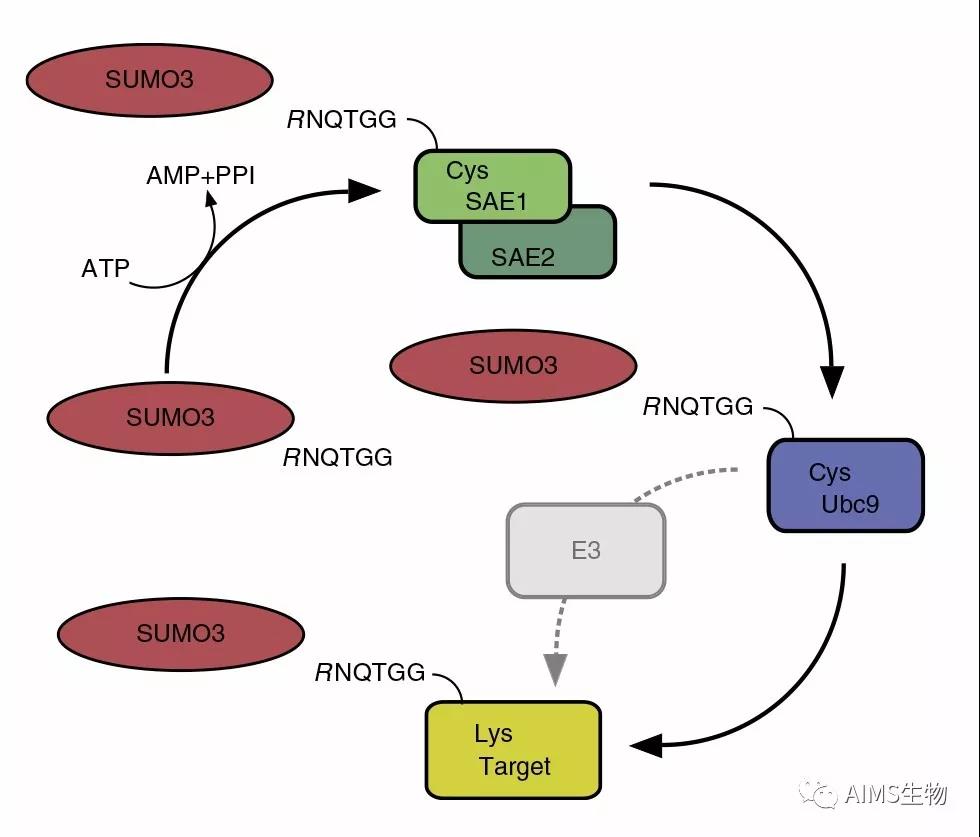
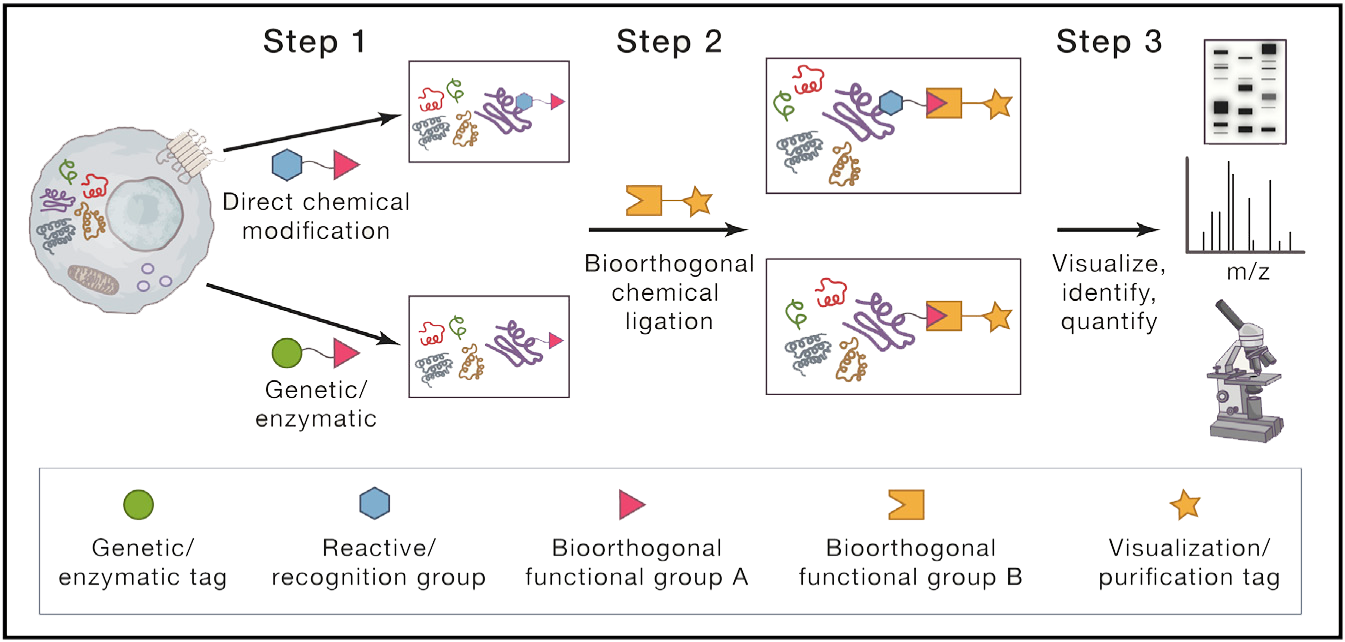
蛋白质棕榈酰化在调控细胞蛋白的运输、稳定性和活性方面发挥着多种作用。点击化学的出现推动了蛋白质棕榈酰化领域向前提供特定的,敏感的,快速的,易于处理的方法研究蛋白质棕榈酰化。本文就近十年来蛋白质棕榈酰化的生物学研究进展进行讨论。特别地,我们讨论了点击化学对实现蛋白质棕榈酰化蛋白质组学方法的影响,揭示蛋白质上新的脂质修饰并阐明其功能,以及发展非放射性生化和酶分析。此外,本综述为利用可点击脂肪酸探针构建和探索蛋白质棕榈酰化的新研究途径提供了背景。
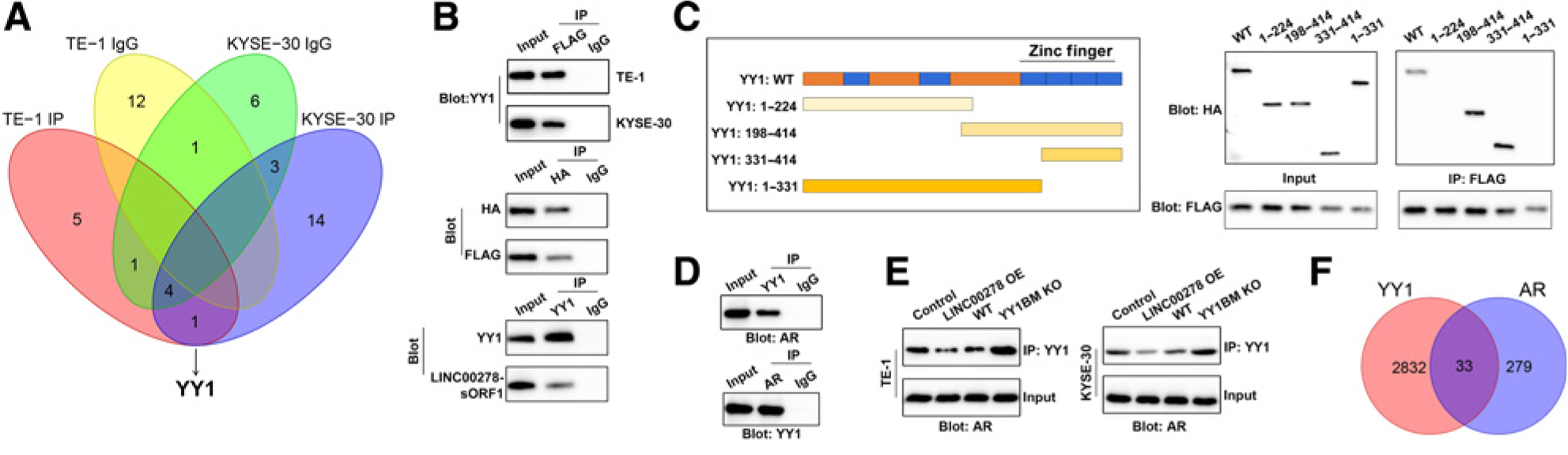
长链非编码rna (lncRNA)已被证实在许多疾病中发挥重要作用,包括食管鳞状细胞癌(ESCC)。 最近的研究报道了一些lncRNA编码功能性微肽。然而,ESCC与lncRNA编码的微肽之间的关系尚不清楚。在该文章中,研究人员鉴定了一个y -连锁lncRNA, LINC00278,它在男性ESCC中下调。LINC00278编码一种阴阳1(YY1)结合微肽,命名为YY1BM。YY1BM参与了ESCC的进展,抑制了YY1与雄激素受体(AR)的相互作用,而雄激素受体(AR)相互作用减弱通过AR信号通路表达eEF2K。YY1BM下调可显著上调eEF2K表达,抑制细胞凋亡,使ESCC细胞更适应营养剥夺。吸烟减少了LINC00278和YY1BM翻译的变形。总之,这些结果提供了一个新的机制联系吸烟和AR信号之间的男性ESCC进展。本文发表在《Cancer Research》,第一作者是苏州大学科研人员,本文质谱部分由Aimsscientific公司协助完成。
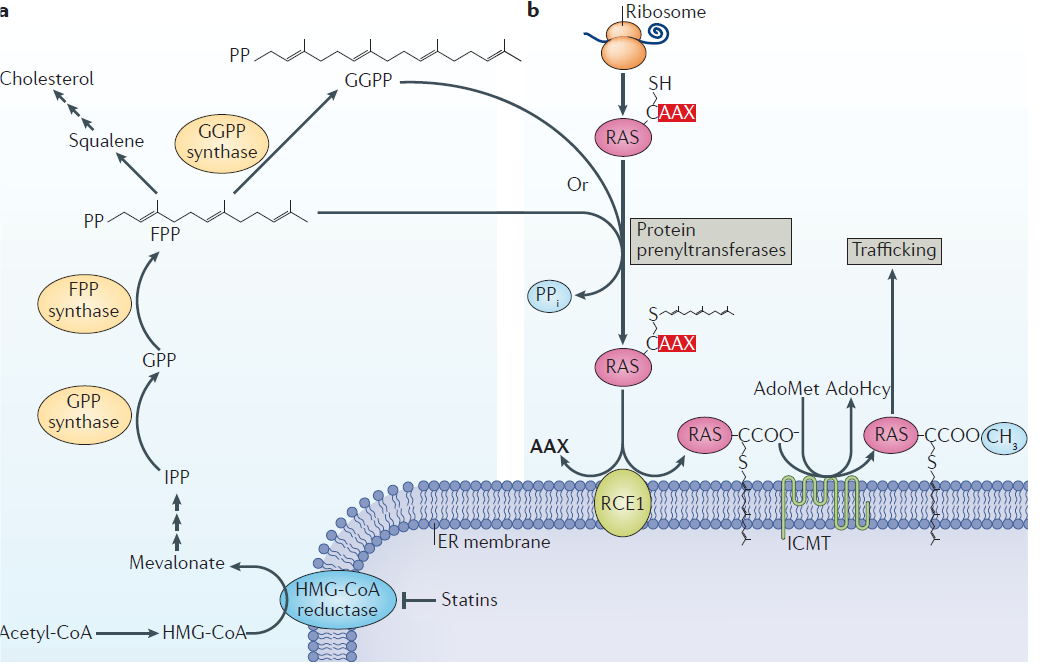
生物体在数十亿年的进化过程中,已经发展出来大量的翻译后修饰种类,以扩大和提高蛋白质功能的复杂性,以应队复杂的外部刺激和内部调整。异戊烯类脂质体对于真核蛋白质的修饰,即所谓的异戊烯修饰,控制着蛋白质定位和活性等一系列在生物调控的重要功能。 异戊烯化修饰蛋白在不同物种之间比较保守,这就突出了脂质修饰途径在生物学和进化过程中的重要性。蛋白异戊烯修饰基于基因抑制以及药理学的抑制剂的相关机制,已经在不同细胞过程和疾病病理学方面深入研究。
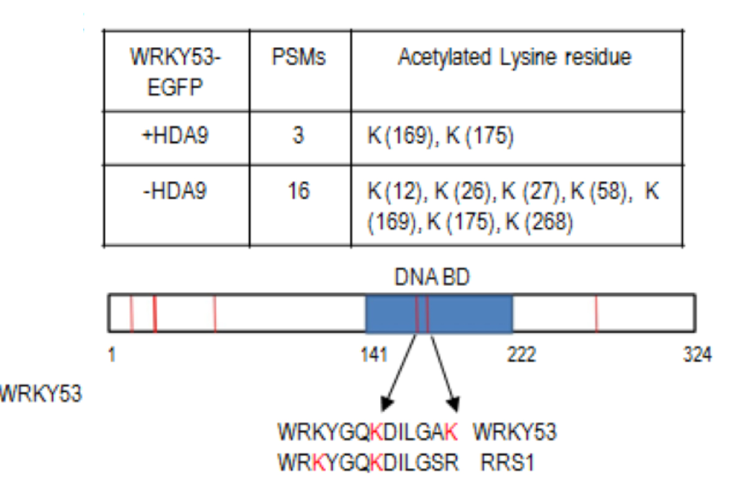
盐引起的非生物胁迫是植物所承受的常见环境胁迫之一,它对植物施加进化压力,反过来植物已经发展处复杂的反应去应对。因此对于盐胁迫反应机制的了解在许多生物技术应用中具有深远的意义,包括提高植物生产力和环境保护。盐胁迫反应的调节是一个受发育信号通路和环境信号通路控制的复杂过程。动态组蛋白乙酰转移酶(HATs)和组蛋白去乙酰化酶之间的平衡(HDACs)控制核小体组蛋白乙酰化水平,影响染色质结构和基因活性。HATs促进基因转录,而HDACs通常与转录抑制和基因沉默有关。
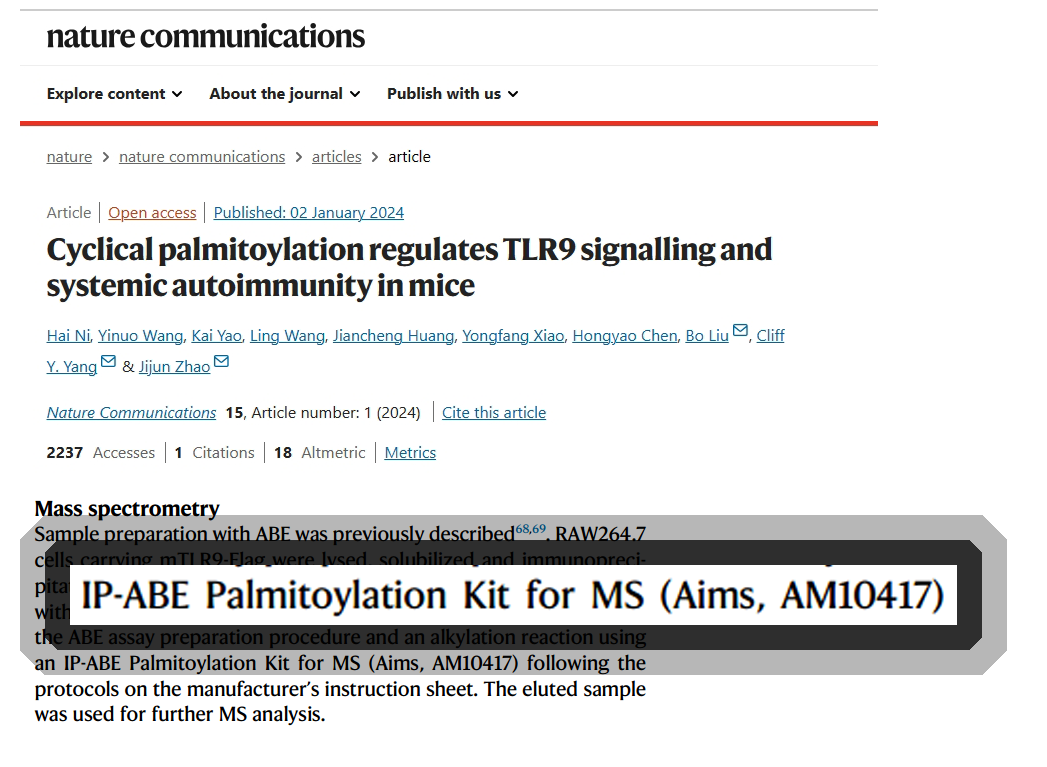
Toll样受体9(TLR9)识别自身DNA并在系统性红斑狼疮(SLE)中发挥复杂的作用。然而,调控内体TLR9反应的分子机制还不清楚。在这里,我们报道了棕榈酰蛋白硫酯酶1(PPT1)通过去除溶酶体TLR9中的s-棕榈酰化来调节全身自身免疫。PPT1促进浆细胞样树突状细胞(pDCs)分泌IFNα和巨噬细胞分泌TNF。遗传缺陷或化学抑制PPT1降低抗核抗体水平,减轻B6Sle1yaa小鼠的肾炎。在健康志愿者和SLE患者中,PPT1抑制剂HDSF可在体外减少IFNα的产生。机制、生化和质谱分析表明,TLR9在C258和C265位点被s-棕榈酰化。此外,蛋白酰基转移酶DHHC3棕榈酰化高尔基体中的TLR9,并调节TLR9转运到核内体。随后PPT1的去棕榈有丝分裂化促进TLR9从UNC93B1释放。我们的研究结果揭示了一个控制TLR9反应和自身免疫的翻译后修饰周期。
本研究中揭示了法尼醇合酶通路(FPP)——胆固醇代谢的中间产物——通过重塑线粒体形态和代谢,在自身免疫性疾病中驱动浆细胞样树突状细胞(mig-DC)功能障碍方面发挥着关键作用。本研究为自身免疫性疾病中 DC 失调的代谢控制提供了见解,并建议了潜在的疾病治疗靶点。